Molecular Self-Assembly in Biomembranes
This page summarizes earlier research work by Martin O. Steinhauser on molecular self-assembly, lipid aggregation, biomembranes, and coarse-grained molecular dynamics simulations. The central question was how amphiphilic molecules organize spontaneously into stable bilayer structures and how this process can be studied by molecular simulation.
Biological membranes are complex supramolecular structures. They separate cellular compartments, regulate transport processes, and provide the physical environment for many biochemical functions. Their stability results from the interplay of hydrophilic head groups, hydrophobic molecular tails, solvent interactions, thermal motion, and collective molecular organization.
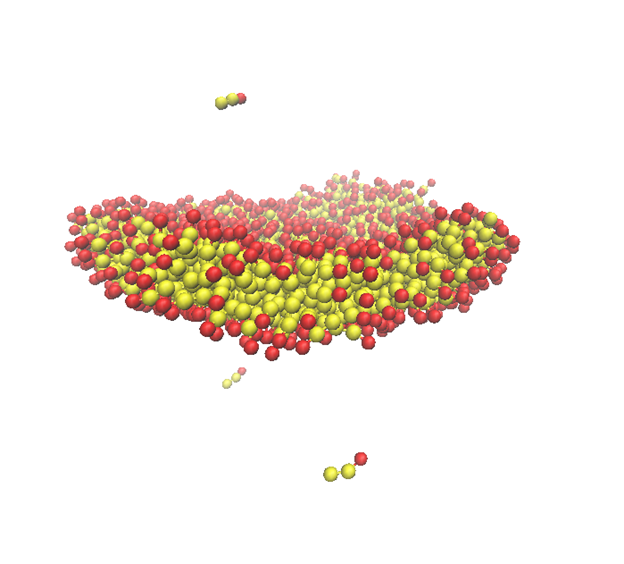
Figure 1 illustrates an early stage of molecular aggregation in a simulated lipid system. Starting from initially disordered molecular configurations, the molecules begin to form clusters that reduce unfavorable contact between hydrophobic parts and the surrounding solvent. Figure 2 shows a more organized bilayer-like structure in which the hydrophilic and hydrophobic molecular regions have separated into a stable membrane configuration.
Concept
The concept of this research line was to use molecular dynamics simulations to study membrane formation as a self-organization process. Instead of prescribing the final bilayer geometry, the simulations allowed molecular aggregates to emerge from the interaction rules of the model. This made it possible to observe how clustering, fusion, reorganization, and bilayer formation develop over time.
A central aspect of the work was the use of coarse-grained models. Such models reduce microscopic molecular complexity while retaining the relevant physical interactions that drive amphiphilic self-assembly. They are especially useful when the processes of interest occur on length and time scales that are difficult to access with fully atomistic simulations.

Applications
The results are relevant for the physical understanding of biomembrane formation, lipid self-assembly, soft matter, and biological interfaces. They also contribute to the broader methodological question of how molecular simulation can be used to study collective processes in complex biological materials.
This earlier work connects molecular modeling, statistical physics, soft-matter science, and biological membrane physics. It also forms part of a broader research background in multiscale modeling, where molecular mechanisms are related to larger-scale structural and functional behavior.
Selected Related Publications
The publications listed below document the molecular simulation methods, coarse-grained models, and self-assembly processes associated with this research line.
Destruction of cancer cells by laser-induced shockwaves: recent developments in experimental treatments and multiscale computer simulations
M.O. Steinhauser, M. Schmidt
SoftMatter, 2014, 10, 4778–4788
Computational Multiscale Modeling of Fluids and Solids
M.O. Steinhauser
Springer, Berlin, Heidelberg, 2017
Multiscale modeling, coarse-graining and shock wave computer simulations in materials science
M.O. Steinhauser
AIMS Materials Science, 4(6): 1319-1357